

CRISPRs-Cas9基因编辑的原理
发布时间:
2019-08-02
1987年日本研究人员在大肠杆菌基因组中发现了成簇的规则间隔回文重复序列CRISPRs(Clustered Regularly Interspaced Palindromic Repeats,CRISPRs)。2005年发现CRISPRs中的许多间隔序列与质粒和病毒具有同源性,最后证实CRISPR-Cas是一种自适应的防御系统,可以使用反义RNA作为过去被入侵的记忆标记,当病毒再次入侵时即可做出快速反应。接下来的研究发现一共存在三种CRISPR-Cas系统类型(I、II和III),它们分别使用不同的分子机制对核酸进行识别和剪切。I型和III型系统使用大量复杂的Cas蛋白作为crRNA介导的DNA剪切,由于其过于复杂,具体应用难度很大。然而,II型系统只需要一个单一的蛋白质Case9,既可以实现基因的剪切,简单方便,后来被证明对基因组工程非常有用,现在被广泛应用于基因编辑和基因治疗。
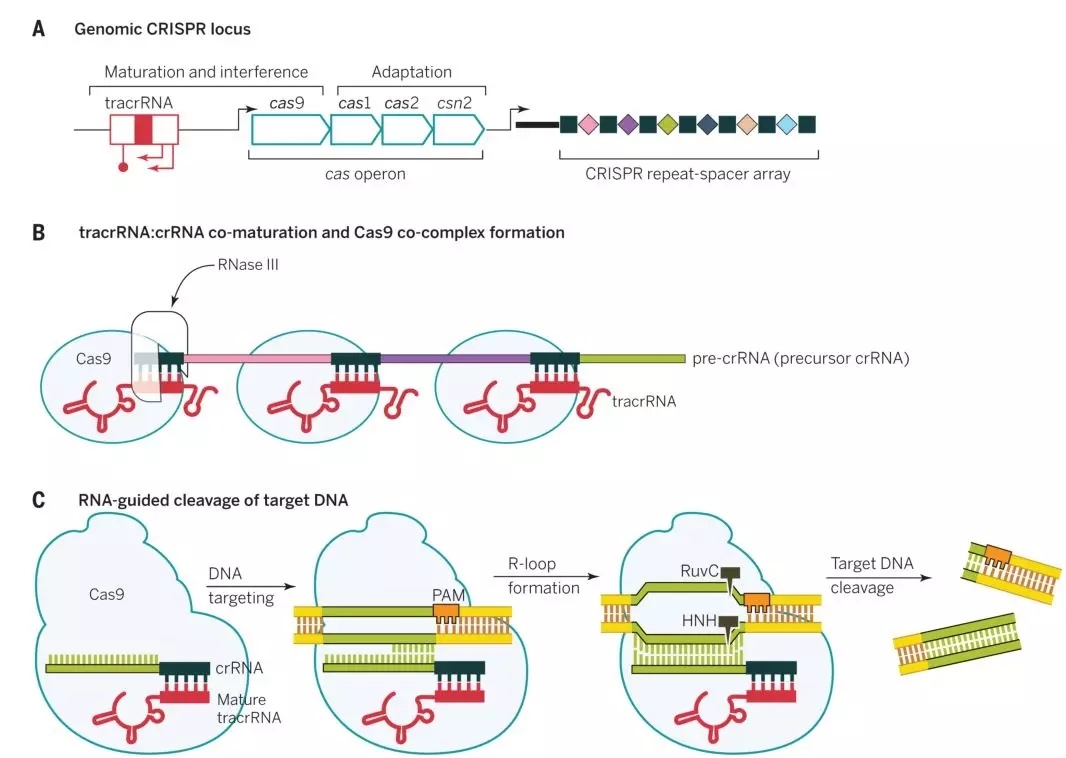
图1 链球菌CRISPRs-Cas9抵抗病毒入侵的机制
上图是链球菌的CRISPRs-Cas9抗病毒防御机制,Cas9是一种RNA引导的DNA核酸内切酶,其需要在crRNA和tracrRNA双RNA的指导下对特定DNA序列进行识别和切割。在CRISPR在基因座里包括tracrRNA、Cas9还有短的重复序列基因(如图1a)。它们发挥作用的具体过程如下:短的重复序列基因转录成pre-crRNA,而tracrRNA对于pre-crRNA的成熟是必须的,Cas9具有核酸剪切能力。它们首先结合形成复合物,在核糖核酸酶Ⅲ作用下进一步成熟(如图1b)。由于crRNA上的20个碱基能与目标DNA的碱基进行互补配对,可将复合物导向目标DNA,随后Cas9对目标基因PAM序列(PAM序列为NGG,N为任意碱基)前的DNA序列进行剪切,导致目标DNA断裂形成DSB(DNA double-stranded break,DBS)使入侵的病毒基因组发生断裂(如图1c),导致病毒死亡,进而抑制病毒感染。人们利用这种机制,通过寻找目的基因特异的PAM序列,将其5‘端前20个碱基一起合成crRNA,然后将tracrRNA和crRNA改造成单链导向的sgRNA(single guide RNA,sgRNA)(图2)。同时共转染Cas9,实现对目的基因的剪切。
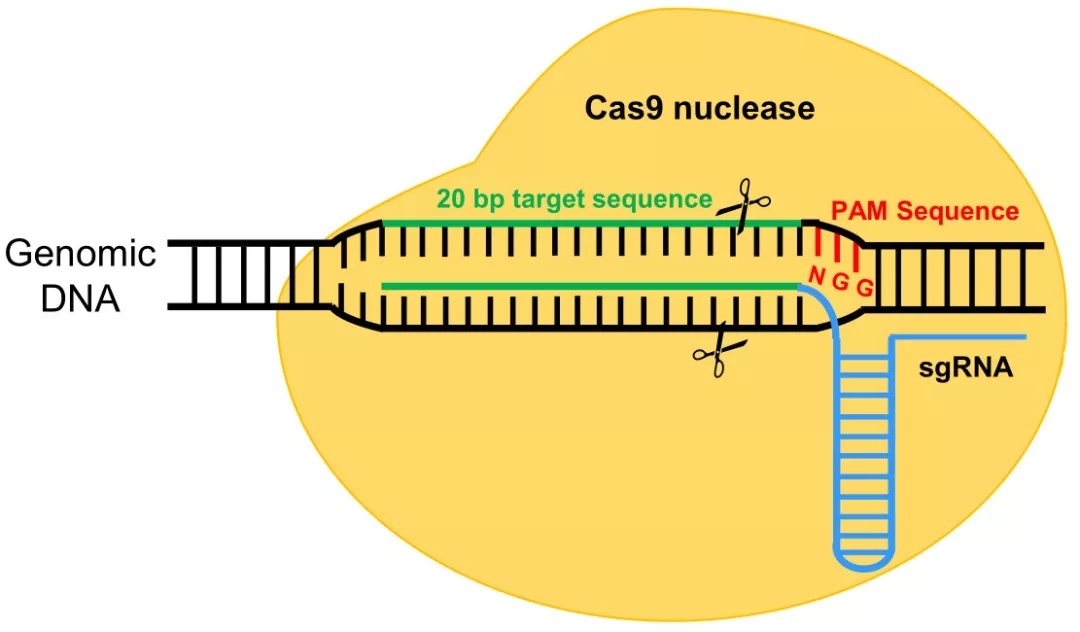
图2 改造后的CRISPs-Cas9系统
然后利用生物体内基因DBS的两种修复方式对目的基因实现编辑。具体两种修复方式包括非同源重组修复(NHEJ)和同源重组修复(HDR)见图3,NHEJ方式的修复会导致目的基因碱基的增加或缺失,导致基因可能发生移码突变,达到基因敲除的目的。而HDR修饰是利用基因同源臂(如等位基因作为模板)可以进行同源重组所以可以进行精确修复,这样人们可以通过外转具有相同同源臂的质粒,在同源臂内插入基因就可以实现对目标基因的替换,进而实现基因编辑。
CRISPs-Cas9技术对基因编辑具有操作简单、成本低、编辑效率较高等优点,已经被广泛应用于疾病、肿瘤等治疗,但是目前还存在脱靶效应、传递效率低、免疫排斥、伦理等问题。
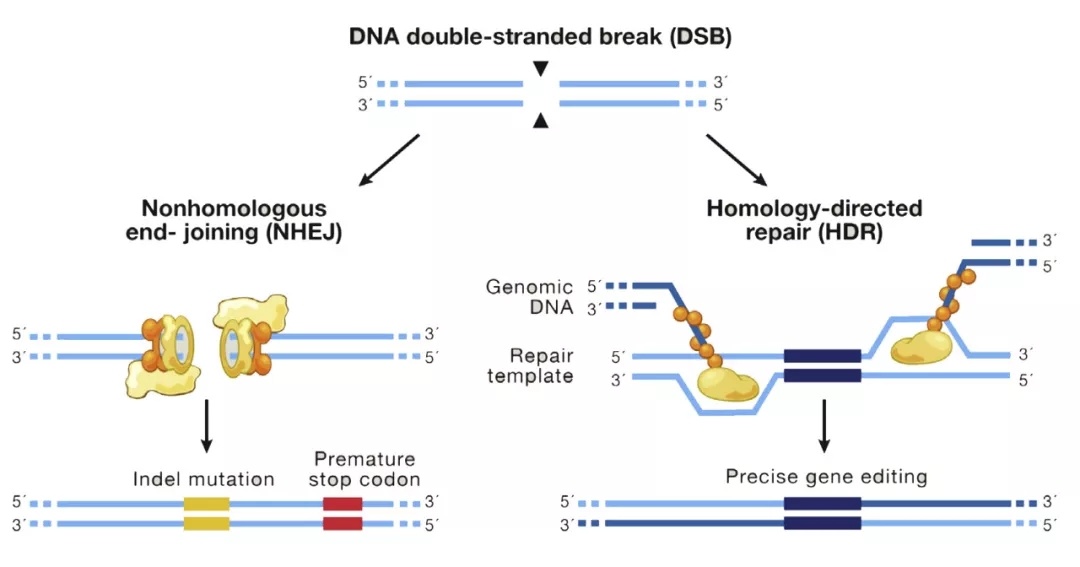
图3 基因组基因断裂修复的两种方式
主要参考文献:Doudna, Jennifer A., and Emmanuelle Charpentier. "The new frontier of genome engineering with CRISPR-Cas9." Science 346.6213 (2014): 1258096.



